
Recently, I was carrying out whole-cell voltage-clamp and LFP recordings with simultaneous optogenetic activation of a channelrhodopsin using blue light. Whole-cell voltage clamp techniques can record the input currents seen by a neuron (previously on this blog [1], [2]); an LFP records the very small synaptic currents in bulk brain tissue (nicely reviewed by Oscar Herreras); and optogentics with genetically encoded rhodopsins can make neurons fire using light pulses.
For the LFP recordings, I used the same glass pipette that I had used before for the whole-cell recording of a nearby neuron. In the LFP, I saw a light-evoked response which I first thought was a rhodopsin-evoked synaptic current. However, it turned out that I could make the same observation when positioning the pipette tip in the bath instead of in the tissue, which meant that this was clearly not a synaptic current, but an artifact. When changing the pipette resistance by gently breaking the pipette tip, the light-evoked voltage remained the same, whereas the evoked currents changed proportionally with the pipette resistance Rp, or more generally with the resistance between the two electrodes:
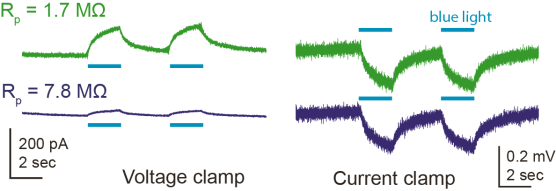
I found out that this sort of artifact has been described in the context of tetrode recordings several years ago by Han et al. (2009; supplementary figure 1) and has been sort of explained with the Becquerel effect (here [update 2022: linked website deleted, therefore the link points to an archived version]), which is better known as the photovoltaic effect. According to Han et al., the effect is stronger for blue light and affects the recorded currents on a slow timescale, such that highpass-filtering of the recorded signal, which is used to detect spikes in tetrode recordings, gets rid of this artifact.
In addition, Han et al. state:
We have not seen the artifact with pulled glass micropipettes (such as previously used in Boyden et al., 2005 and Han and Boyden, 2007, or in the mouse recordings described below). Thus, for recordings of local field potentials and other slow signals of importance for neuroscience, hollow glass electrodes may prove useful.
Contrary to this suggestion, my above measurements indicate that using a glass electrode does not or not always get rid of the artifact. To better understand this artifact, I checked whether it was mediated by the chloride silver electrode in the glass pipette or rather by the ground electrode, and found that both contributed more or less equally to the artifact in this experiment. Protection of the electrode by some sort of cover reduced the magnitude of the artifact.
What does this mean for whole-cell or LFP recordings using a glass pipette? For whole-cell recordings, the resistance between the two electrodes is much larger than for the two traces shown in the plots above, typically between 50 and 2000 MΩ. This reduces the artifact-induced current recorded in voltage-clamp to something less than 5 pA for 50 MΩ cells, and much less for neurons with higher membrane resistance. In most cases, this is negligible.
For glass pipette-based LFP recordings or loose seal cell-attached recordings, however, the light-induced voltage change (few hundred μV, as shown above) is of the same magnitude as a strong LFP signal (see for example figure 1 in Friedrich et al., 2004). Therefore, in order to measure a LFP signal in response to blue light-activated rhodopsins, one needs to take into account the artifacts induced by the photovoltaic effect. This can for example be done by measuring the light-evoked voltage change with the glass pipette both in the tissue and in the bath and subtracting the latter measurement from the previous one on a pipette-by-pipette basis.
I would also be curious about other reports (if there are any) on light-induced artifacts with recording electrodes and under which circumstances (if there are any) they might play a non-negligible role.

Pingback: Whole-cell patch clamp, part 1: introductory reading | A blog about neurophysiology
Pingback: Whole-cell patch clamp, part 4: look and feel | A blog about neurophysiology