
To record the activity from a population of neurons, calcium imaging and extracellular recordings with small electrodes are the two most widely used methods that are still able to disentangle the contributions from single units. Here, I would like to briefly mention two papers that try to connect these two approaches by comparing them more or less directly.
- Wei et al., A comparison of neuronal population dynamics measured with calcium imaging and electrophysiology, bioRxiv, 2019
[Update, 2020-09-15: The paper just came out in PLoS Comp Biology just one day after the blog post!] - Siegle, Ledochowitsch et al., Reconciling functional differences in populations of neurons recorded with two-photon imaging and electrophysiology, bioRxiv, 2020
Both papers compare calcium imaging datasets and extracellular ephys datasets, try to connect the results and point out the difficulties in reconciling the approaches.
Wei et al. use datasets recorded in mouse anterio-lateral motor cortex (ALM). They first focus on approaches to reconstruct spike rates from calcium imaging data (deconvolution) and find several limitations of this approach. On the other hand, they find that a forward model that transforms spiking activity to calcium fluorescence data can reconcile most of these differences.
The authors also provide a user-friendly website which can be used to explore the transformations between ephys and imaging data (also including datasets with simultaneous ephys-imaging): http://im-phys.org. (Understanding the figures of the paper is however quite useful before exploring the website.)
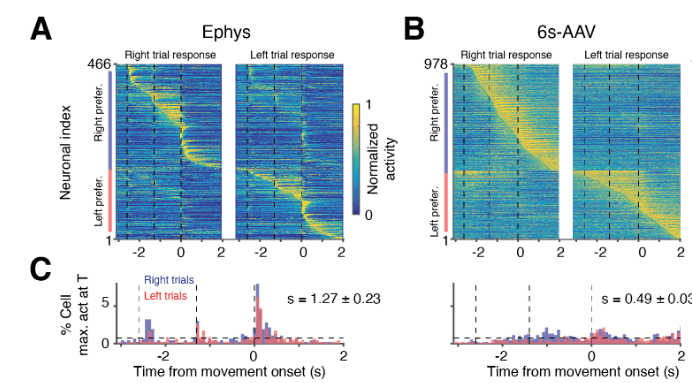
A large part of the paper focuses on high-level analyses (principal component analysis, decoding of behavior and decision). I would take it as an educational tale of caution which highlights wrong conclusions that could be made based on standard analyses. For example, the slow and variable decay times of calcium imaging data can lead to a dispersion of peak activity that was absent in the ephys data. This dispersion can smear out clearly timed activations of neuronal population into something more similar to a sequence (see Figure 7, of which I have pasted an excerpt above).
Siegle, Ledochowitsch et al. from the Allen Institute, rather than investigating the effects on higher-level population analyses, focus their attention on the effects seen in single neurons. When comparing a calcium imaging and an ephys dataset recorded in the same brain region (visual cortex V1) in mice that do the same standardized tasks, what differences can be seen in the firing properties of single neurons?
Due to the high standardization requirements at the Allen Institute, their datasets are probably uniquely qualified to be the basis for such a comparison. Interestingly, they find a couple of clear differences. For example, extracellular ephys data suggest typical firing rates of around 3 Hz (see Figure 2A), which is almost an order of magnitude higher than what has been recorded and estimated from calcium imaging data (see also Figure 7 in our preprint, which estimates spike rates of the same dataset).
The authors go to great lengths to use forward transformations (similar to Wei et al.) in order to reconcile differences seen for various response metrics (responsiveness, tuning preference, selectivity). However, their conclusion seemed to me quite a bit less optimistic compared to the Wei et al. paper. The authors go into more detail when discussing the potential reasons for the discrepancies, and focus on the recording methods themselves rather than on methods to transform between them. In particular their analysis of inter-spike-interval (ISI) violations in ephys recordings (which indicate that spikes from different neurons contaminate the recording of the neuron of interest) was, in my opinion, particularly interesting and convincing. I also really recommend to anybody the last paragraph of their discussion, from which I only want to cite their note of caution about extracellular ephys recordings:
From this study, we have learned that extracellular electrophysiology overestimates the fraction of neurons that elevate their activity in response to visual stimuli, in a manner that is consistent with the effects of selection bias and contamination. – Siegle, Ledochowitsch et al.
One of the reasons why I am writing about these two studies is that I have been working at the interface of calcium imaging and ephys myself, addressing the question, How much information about spike rates can we get out of calcium imaging data? Wei et al. and Siegle, Ledochowitsch et al. take a slightly broader perspective. And, in some way, they show how hard it is to reconcile two (methodological) perspectives on the same phenomenon. (I noticed this in my PhD lab as well, when it came to reconciling results from EM reconstructions of neuronal anatomy and calcium measurements of the same neurons.) Since almost any method in systems neuroscience is technically challenging, we often have in a single lab only a single perspective of a phenomenon, and I think it’s important to always be aware that the conclusions drawn from this perspective might be strongly biased.
In general, calcium imaging and extracellular ephys are extremely valuable tools to observe the living brain, and we better do anything to understand the properties and limitations of these tools. These studies sometimes might feel a bit like negative results and therefore not very attractive to those who want to advance neuroscience, and I therefore understand why not many are willing to undertake these projects. So I am glad to be able to highlight these two publications here.

Pingback: Temporal dispersion of spike rates from deconvolved calcium imaging data with Cascade | A blog about neurophysiology