
It requires more than a quick look at the abstract and the figures to fully understand a research paper and its limitations. One way to go there is to write a summary or critical review of a paper. In a contribution to (informal) post-publication review, I will select neuroscience papers that are, in my opinion, worth the time needed to write up a review. On L5 apical dendrites [2]; on simulations of calcium imaging datasets [3].
Target audience: Theoretical neuroscientists working on balanced networks and the role of inhibition; people working on the CA3 to CA1 feedforward circuit in hippocampus; people working on rules governing the connectivity of inhibitory interneurons; people interested in codes dependent on precise spike timing.
Why I chose this paper: I have worked on a similar topic at the interface of theoretical and experimental neuroscience, the so-called ‘precise balance of excitation and inhibition’ (Rupprecht and Friedrich, Neuron (2018)). Of course I’m interested what other experimental studies have found out about this important, although not very easily accessible concept.
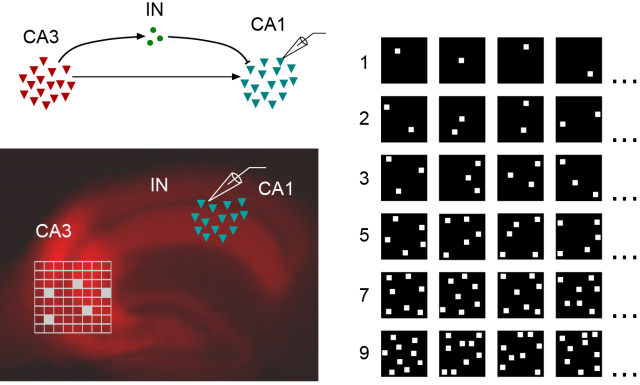
From Bhatia et al., eLife (2019), used under CC BY 4.0 license (modified from Fig. 1).
Summary: The authors use a slice preparation to dissect the feedforward circuit from CA3 to CA1 in the mouse hippocampus (figure above; left part).
Two main findings.
The first finding (‘detailed balance’): They stimulate CA3 that expresses channelrhodopsin with an artificial set of patterned light stimuli (figure above; lower left and right parts). Then, they use whole-cell recordings to measure inhibitory and excitatory currents in single neurons in CA1. They find that even for very weak stimuli that presumably elicit few or even single action potentials only in CA3, an inhibitory current is evoked in CA1. Moreover, and even more interesting, the size of this inhibitory current matches the excitatory current not only on average, but also for individual stimuli. This feature has been termed ‘detailed balance’ by theoretical neuroscientists.
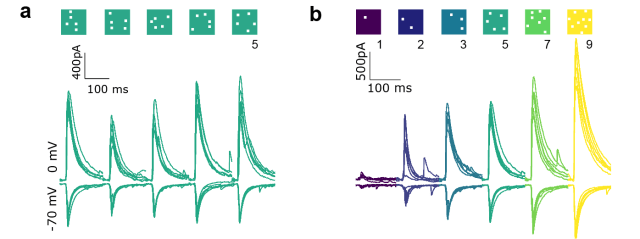
From Bhatia et al., eLife (2019), used under CC BY 4.0 license (modified from Fig. 2).
The second main finding: The balanced inputs as recorded with voltage-clamp end up, however, in a non-linear integration as observed with current-clamp recordings. In particular, the authors observe a supra-linear relationship which they describe as ‘subthreshold divisive normalization’. In addition, they notice that inhibition kicks in faster in the CA3 neuron when the excitatory (and inhibitory) inputs to the CA3 neuron are stronger. They use this finding and simulations to support the idea that the reduction of inhibitory delays is the underlying cause of the observed divisive normalization.
In the following, I will only discuss the first finding. The second finding is already broadly discussed in the paper and also by the reviews (which are partly accessible, thanks to eLife!). In general, I find the first finding (detailed balance) much more interesting, whereas the main focus of the paper is on the second finding.
Consequences of a detailed balance: A detailed balance in this feedforward path from CA3 to CA1 is something unexpected, because it would require very specific feedforward connectivity of interneurons matching the feedforward connectivity of excitatory connections. Importantly, the authors put this in the context of previous work on CA1, which found that almost any CA1 cell can be converted into a place cell for any spatial location:
“Precisely balanced networks, with all input subsets balanced, are well suited for input gating (Barron et al., 2017; Hennequin et al., 2017). The finding that most silent CA1 cells can be converted to place cells for arbitrary locations predicts the existence of an input gating mechanism (Lee et al., 2012), but the nature of this mechanism remains unknown. One prediction of precise balance is that inputs for multiple potential place fields may be balanced, and hence place field activity is gated ‘off’. Evoked depolarizations (Lee et al., 2012) or dendritic plateau potentials (Bittner et al., 2015; Bittner et al., 2017), which potentiate the subset of active synapses, that is, change the I/E ratio (Grienberger et al., 2017), can flip the gate ‘on’, thereby converting a silent cell to a place cell for that specific place field”, write Bhatia et al., eLife (2019).
The devil’s advocate: Detailed balanced means, as mentioned before, that the size of an inhibitory current matches the excitatory current not only on average, but for individual stimuli. It is important, as I have described in my own experimental work (Rupprecht et al., Neuron (2018)), that this statement of a detailed balance can only be made if the different individual stimuli are equally strong.
It is therefore not a good idea to assess detailed balance by pooling over all stimuli with 1, 3, 5, 7 and 9 squares of optogenetic activation. Instead, one should compare within a set of stimuli with e.g. 1 square. This is done, as far as I understand it, only in Fig. 2h (example shown for a single neuron) and, based on a current-clamp dataset, in Fig. 2/Supplement 2e-f. It would have been interesting to analyze this aspect in more depth.
More importantly, even when analyzing e.g. only stimuli that all of them correspond to the stimulation of one “square” with photoactivation, this does not mean that all those stimuli are equally strong – which seems to be asssumed in the modeling part (Fig. 2/Supplement 2i,k). As I understand it, the most parsimonious explanation of the data would not be a detailed balance of excitatory and inhibitory inputs in this feedforward circuit, but a distribution of activation strengths for each stimulus set.
For example, stimulation of grid position 3 may overall result in 17 action potentials in CA3, whereas stimulation of grid position 7 may result in 55 action potentials and grid position 9 in zero action potentials. Feedforward inhibition could then simply integrate all action potentials in CA3 and send out a global signal proportional to this number. Indeed, even a single interneuron could, in theory, do this job. Therefore, it seems that the results can be explained by simple and plausible assumptions that do not require detailed balance. Altogether, I think that the paper provides some evidence for a detailed balance in the CA3 to CA1 feedforward circuit, but given the unexpectedness of this result, the evidence is a bit too weak to make me believe the conclusion entirely.
To fully test for a detailed balance, one could try to further analyze the existing data. However, a more direct way to test it would be much more convincing. This could be done by stimulation of CA3 and recording currents in CA1 as before, but at the same time recording the precise number of action potentials elicited in CA3 neurons. The best and maybe only reliable way to do so would be to perform paired recordings, with one (or more) neuron(s) patched in CA3 that are triggered to fire a given number of action potentials, while recording from one (or more) neuron(s) in CA1 in voltage-clamp. This would require considerably more work and result in much lower throughput than the opto-activation experiments, but would allow to address the question of a detailed balance directly and with higher precision.
Conclusion: Overall, I liked the study because it uses experiments to address an important question from theoretical neuroscience. The finding of a balance of excitation and inhibition in CA1 even for (presumably) very few spikes elicited in CA3 is an interesting and unexpected finding which is clearly supported by the data. I’m a bit less convinced about the evidence that this balance is also detailed. However, this does not affect anything of the remaining paper (which constitutes its main part). This main part investigates the supra-linear summation of inputs and the information contained in the timing of inputs (which I do not want to discuss here in any detail).
More context: What is detailed and precise balance? Hennequin et al., Annual Review of Neuroscience (2017). – A broadly accessible explanation of the supra-linear summation presented in the paper: NCBS News.

Pingback: Post-publication review: Somato-dendritic coupling of L5 neurons in V1 | A blog about neurophysiology
Pingback: Review: An artificial ground truth for calcium imaging | A blog about neurophysiology